Odborné články
Fosfolipidové mastné kyseliny (PLFA) jako indikátory změn mikrobiálních půdních složek
Summary:
The analysis of soil microbial phospholipid fatty acids proved the assumption,
that these substances are suitable as bioindicators reacting to the natural and
anthropogenous stress factors. Phospholipids are the basic construction
component of cell membranes and are exposed to the influence of intracellular
and extracellular environment. Under the thumb of the environment in which the
microorganisms lives, the composition of the phospholipids and the structures of
the microbial communities change.
Mastné kyseliny jsou hlavní složkou buněčných membrán živých buněk, zde jsou esterifikované především trojsytným alkoholem glycerolem, kdy jsou dvě OH skupiny navázány na mastné kyseliny což tvoří takzvanou hydrofobní část glykolipidu a na poslední funkční OH skupinu glycerolu je pak navázána kyselina fosforečná. Kyselina fosforečná pak tvoří hydrofilní skupinu. Tyto fosfolipidy vytváří dvojitou buněčnou membránu. Na následujícím schématu je patrné rozdělení fosfolipidových mastných kyselin, kdy esterifikované mastné kyseliny tvoří 60 až 90 % mastných kyselin obsažených v buněčné membráně. Další skupinou tvořící buněčnou membránu jsou neesterifikované mastné kyseliny (10 až 40 %). Esterifikované kyseliny se dále rozdělují na esterifikované nesubstituované a hydroxy-substituované mastné kyseliny, nesubstituované se pak dále dělí na nasycené a na nenasycené mono a poly- nenasycené. Nasycené kyseliny vytvářejí dvě další skupiny s rozvětveným a nerozvětveným řetězcem. Neesterifikované mastné kyseliny se rovněž rozdělují na nesubstituované a hydroxy-substituované. Hydroxy-substituované neesterifikované mastné kyseliny se nacházejí v lipopolysacharidech tvořících buněčnou stěnu Gram negativních bakterií.
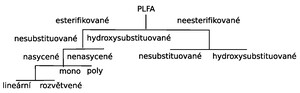
Každá skupina těchto PLFA nebo jejich specifické poměry jsou charakteristické pro některou ze skupin mikroorganismů běžně se nacházejících v mikrobiálně osídlených substrátech. Jakékoliv změny a to jak přirozené tak antropogenně podmíněné mohou vyvolat změnu poměru jejich zastoupení. Jsou-li známy faktory, které změny vyvolaly je možné z nerovnováhy v optimálním poměru skupin mikroorganizmů předpokládat obdobný vliv xenobiotik nebo antropogenních zásahů jako na půdách testovacích.
Rychlá metoda byla původně vyvinuta pro hodnocení čistých bakteriálních izolátů a bakteriálních směsí. Naším cílem bylo ověřit zda a v jaké míře lze tuto metodu využít i pro analýzu nativního půdního vzorku. K nativnímu vzorku půdy o hmotnosti 1 g bylo přidáno 10 ml roztoku sestávajícího ze 45 g NaOH, 150 ml metanolu doplněného 150 ml demineralizované vody. Tato směs byla zahřívána ve vodní lázni při 100°C po dobu 30 minut, tak aby veškeré bakterie hydrolyzovaly a mastné kyseliny byly převedeny na sodné soli. Po ochlazení preparátu ve studené vodě byla přidáno 20 ml roztoku sestávajícího z 325 ml 6N HCl a 275 ml metanolu. Tento roztok byl intenzivně protřepán a inkubován po dobu 5 minut při 80°C. Tento krok převedl sodné soli kyselin na metylestery. Pak byly vzorky opět ochlazeny na pokojovou teplotu ve vodní lázni. Pro extrakci esterů mastných kyselin byl použit roztok sestávající z 200 ml hexanu a 200 ml methyl-terc-butyl-éteru. Pro odstranění volných mastných kyselin byl k této fázi přidán roztok sestávající z 10,8 g NaOH a 240 g NaCl v 900 ml demineralizované vody. A opětovně bylo prováděno intenzivní třepání po dobu 5 minut. Horní organická fáze byla po odstátí odsáta a aplikována do plynového chromatografu Labio.
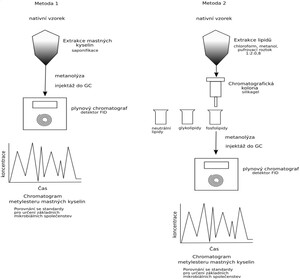
Experiment
Pro experimenty byly připraveny dva vzorky půd stejného složení odebrané z lokality Chomutov. Jeden vzorek byl uměle kontaminován solemi zinku (ZnSO4 .7H2O), kadmia (CdCl2), olova Pb(NO3)2 a mědi CuSO4.5H2O, tak aby došlo ve všech případech k překročení limitních koncentrací těchto látek v půdách podle vyhlášky MŽP číslo 13/1994 Sb. Koncentrace vybraných prvků v tomto vzorku jsou uvedeny v tabulce číslo jedna.
Tabulka číslo 1: Koncentrace prvků v uměle kontaminovaném vzorku mg kg-1
| prvek | obsah v mg kg-1 |
|---|---|
| Cd | 4,8 |
| Pb | 1864 |
| Zn | 3724 |
| Cu | 760 |
Druhý vzorek pak byl nekontaminovaný a obsahy vybraných rizikových prvků odpovídaly standardními pozaďovým hodnotám. V následující tabulce číslo dvě jsou uvedeny obsahy vybraných rizikových prvků v nekontaminovaném vzorku.
Tabulka číslo 2: Koncentrace prvků v původním nekontaminovaném vzorku mg kg-1
| prvek | obsah v mg kg-1 |
|---|---|
| Cd | 0,4 |
| Pb | 20 |
| Zn | 60 |
| Cu | 40,8 |
Pro zachování stejných podmínek byly vzorky umístěny do termostatu, kde byla udržována teplota 28°C a vlhkost 95%. Vzorky byly inkubovány v termostatu po dobu 14 dnů. Po této době byly vzorky podrobeny analýze fosfolipidových mastných kyselin rychlou metodou vyvinutou pro softwarové vybavení Sherlock fy MIDI Inc. USA.
Pro vyhodnocení výsledků byla použita metoda analýzy PLFA při které se vyhodnocují specifické mastné kyseliny charakteristické pro skupiny půdních mikroorganizmů. V následující tabulce jsou popsány skupiny bakterií a odpovídající fosfolipidové frakce, které jsou podle vyjmenovaných autorů charakteristické pro danou skupinu.
Tabulka číslo 3: Fosfolipidové frakce extrahované z půdy a jejich příslušnost ke skupině mikroorganismů a prvoků
| Skupina mikroorganismů | PLFA markery | Literární zdroje |
|---|---|---|
| Gram -negativní | cy17:0, cy19:0,16:1ω 5c, 18:1ω 9c, 15:1ω 4c, 18:1 ω 7c, 17:1ω 9 c | O’Leary and Wilkinson, 1988; Zelles et al., 1994; White et al.,1996; Zelles, 1997 |
| Gram -pozitivní | i14:0; i15:0; a15:0; i16:0; i17:0; a17:0 | |
| Houby | 18:2ω 6c | Frostegard et al., 1993; Zelles, 1997 |
| Aktinomycety | 10Me16:0; 10Me17:0; 10Me18:0 | Kroppenstedt, 1985; Brennan, 1988; Zelles et al., 1994 |
Vysvětlivky: cy cyklopropan; ω umístění dvojné vazby u omega konce kyseliny; xx:y počet uhlíků v řetězci : počet dvojných vazeb; i iso konformace; a anteiso konformace; c cis konformace; t trans konformace; Me umístění metylové skupiny
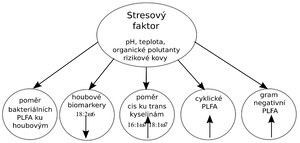
První výsledky jsou patrné z výše uvedené tabulky číslo čtyři. V kontaminované půdě inkubované po dobu 14 dní v termostatu a v původním vzorku došlo ke změně spektra mikrobiálních společenství. Zatímco v kontrolní půdě bylo zastoupení G-negativních a G-pozitivních bakterií charakterizovaných spektrem PLF patrným z tabulky číslo tři takřka totožné, došlo u kontaminované půdy k úbytku G-pozitivních bakterií. Rovněž byl u kontaminované půdy patrný pokles fosfolipidů extrahovaných z hub. Pro průkaznost vlivu stresových faktorů lze využít poměr nasycených k nenasyceným kyselinám (12:0 + 13:0 +14:0 +15:0 +16:0 +17:0 +18:0 +20:0 / 14:1ω 5c +15:1ω 6c + 16:1ω 7c + 16:1ω 5c + 17:1ω 9c +18:1ω 9c +18:1ω 7c). Tyto poměry jsou dle těchto autorů (Knivett and Cullen, 1965; Kieft et al., 1997; Bossio and Scow, 1998 ) vhodné pro vyjádření účinku stresových faktorů. Dále se vlivem biotických a abiotických stresů mění poměr mezi houbovými kyselinami (18:2ω 6c) a bakteriálními kyselinami ( i14:0; i15:0; a15:0; i16:0; i17:0; a17:0 16:1ω 9c, 18:1ω 9c, 15:1ω 4c, 18:1ω7c, 17:1ω 9 c) Rovněž se podle různých autorů předpokládá, že v půdách kontaminovaných rizikovými látkami dochází ke snížení počtu Gram-pozitivních bakterií (Frostegard et al., 1993), zatímco počet Gram-negativních zůstává nezměněn. Zvýšená odolnost Gram-negativních bakteriálních kmenů se vysvětluje stabilnějšími „cyklo“ mastnými kyselinami a přítomností lipopolysacharidů v buněčných membránách (Guckert et al 1986). V následující tabulce číslo 4 jsou uvedeny výsledky získané analýzou extraktu půdy s pozaďovými hodnotami a půdy kontaminované vybranými rizikovými látkami.
Tabulka číslo 4: Výsledky získané analýzou chromatogramu PLFA kontaminované a kontrolní půdy v % celkového množství metylesterů mastných kyselin ve vzorku
| Zemina | Vzorek | G-pozitivní bakterie | G-negativní bakterie | Aktinomycety | Houby | Houby / bakterie | Nasycené / nenasycené |
|---|---|---|---|---|---|---|---|
| Kontamin | A | 16 | 20 | 12 | 1,1 | 0,03 | 1,25 |
| B | 19 | 21 | 17 | 0,9 | 0,022 | 1,10 | |
| C | 18 | 21 | 13 | 1 | 0,026 | 1,17 | |
| Kontrola | A | 25 | 21 | 11 | 1,6 | 0,035 | 0,84 |
| B | 23 | 19 | 12 | 1,8 | 0,043 | 0,82 | |
| C | 21 | 22 | 14 | 1,5 | 0,035 | 1,04 |
Analýza fosfolipidových mastných kyselin půdních mikroorganizmů potvrdila, že jsou tyto látky vhodné jako bioindikátory, reagující na stresové faktory a to jak přírodní tak antropogenní. Fosfolipidy jsou základní stavební složkou mikrobiálních buněčných membrán a jsou vystaveny vlivu intracelulárního i extracelulárního prostředí. To má za následek, že vlivem prostředí v němž mikroorganizmy žijí dochází ke změnám ve složení samotných fosfolipidů, ale i ke změnám struktury samotných mikrobiálních společenstev. Pro hodnocení vlivu faktorů ovlivňujících kvalitu i kvantitu fosfolipidových mastných kyselin je možné použít následující parametry, které se vlivem stresoru v mikrobiální populaci mění a jsou měřitelné.
Tyto výsledky byly realizovány v rámci etapy 4 „Techologie zpracování a kompostování biodegradabilních odpadů a využití kompostů pro zlepšení bilance organických látek a sekvestrace C do půdy“ výzkumného záměru 0002700601 „Principy vytváření, kalibrace a validace trvale udržitelných a produktivních systémů hospodaření na půdě“.
TweetČlánek: Tisknout s obrázky | Tisknout bez obrázků | Poslat e-mailem
Související články:
Obsah těkavých mastných kyselin ve vstupních surovinách pro přípravu kompostu
Evropské priority výzkumu nakládání s organickými odpady v letech 2007 až 2013
Zobrazit ostatní články v kategorii Obnovitelné zdroje energie
Datum uveřejnění: 6.3.2006
Poslední změna: 5.3.2006
Počet shlédnutí: 18064
Citace tohoto článku:
HONZÍK, Roman, SLEJŠKA, Antonín: Fosfolipidové mastné kyseliny (PLFA) jako indikátory změn mikrobiálních půdních složek. Biom.cz [online]. 2006-03-06 [cit. 2026-07-16]. Dostupné z WWW: <https://biom.cz/cz-spalovani-biomasy-pelety-a-brikety/odborne-clanky/fosfolipidove-mastne-kyseliny-plfa-jako-indikatory-zmen-mikrobialnich-pudnich-slozek>. ISSN: 1801-2655.
Komentáře:

|

|

|

|

|